 PDF(6450 KB)
PDF(6450 KB)

北方白菜型冬油菜BrFLC家族的全基因组鉴定、克隆及表达分析
姚彦林, 马骊, 刘丽君, 李学才, 李鹏, 王旺田, 蒲媛媛, 牛早霞, 徐芳, 孙万仓, 武军艳
中国油料作物学报 ›› 2023, Vol. 45 ›› Issue (1) : 83-94.
PDF(6450 KB)
PDF(6450 KB)
北方白菜型冬油菜BrFLC家族的全基因组鉴定、克隆及表达分析
Genome-wide identification, cloning and analysis of BrFLC family in northern winter rapeseed (Brassica rapa)
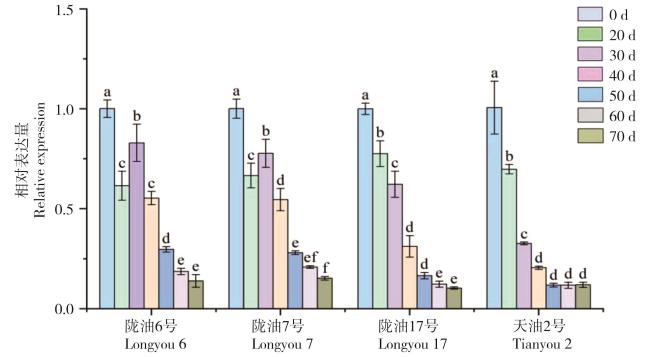
春化过程是冬性植物抽薹开花前的主要阶段。FLC(FLOWERING LOCUS C)基因在春化过程中可以整合上游信号调控植物开花。为了解白菜型冬油菜中FLC基因的特征及其春化功能,以6个拟南芥FLC基因为索引从白菜型冬油菜基因组中鉴定得到10个BrFLC基因,其中8个均含有7个外显子。10个BrFLC分布于4条染色体上,存在明显的基因复制现象。进化及保守结构分析显示聚为一类的成员蛋白质保守Motif分布相似。基因上游1500 bp的启动子区域均含有光应答元件,其中BrFLC4、BrFLC5和BrFLC6含低温响应元件,BrFLC3、BrFLC4、BrFLC7、BrFLC9含激素响应元件。比对发现,位于A02上的BrFLC6基因序列与大白菜中已注释的Bra031886差异性较大。克隆到陇油6号、陇油17、天油2号和天祝小油菜中的BrFLC6基因,序列相似性达99.5%。分析其中序列差异最大的BrFLC6基因,qRT-PCR检测发现它可在根、茎、叶、花蕾及花中表达;春化处理后则表达下调,不同品种间下调幅度有差异。结合春化率的观察,认为春化过程抑制了BrFLC6的表达,且基因表达水平与白菜型冬油菜通过春化所需的时间有关。
Vernalization is the main stage of bolting and flowering of winter plants, FLC (FLOWERING LOCUS C) integrates upstream signals to regulate flowering during vernalization. To understand the characteristics of FLC genes in winter rapeseeds (B. rapa) and its function in vernalization, 10 BrFLCs were identified from winter rapeseed genome using 6 Arabidopsis FLC genes as queries, 8 of which contained 7 exons. 10 BrFLCs are distributed on 4 chromosomes with obvious gene duplications. Evolutionary and conserved structure analysis showed that members in the same class were clustered with similar protein conserved motif distributions. The 1500 bp upstream promoter region of all the genes contained light response elements, among which BrFLC4, BrFLC5 and BrFLC6 contained low temperature response elements, and BrFLC3, BrFLC4, BrFLC7 and BrFLC9 contained hormone response elements. BrFLC6 gene sequence located on A02, it was quite different from the annotated Bra031886 in B. rapa. Thus weconsequently cloned the BrFLC6 genes from Longyou 6, Longyou 17, Tianyou 2, and Tianzhu small rape, and found that their sequence alignment similarity reached 99.5%. By analyse the BrFLC6 gene with the greatest sequence difference, qRT-PCR detection showed that could be expressed in roots, stems, leaves, flower buds and flowers. BrFLC6 showed a down-regulation trend after vernalization, but the down-regulation extent was different among cultivars. Combined with the vernalization rate, it indicated that vernalization process might inhibited BrFLC6 expression, and its expression level was related to the time required by winter rapeseeds to pass through vernalization.
白菜型冬油菜 / FLC转录因子 / 生物信息学分析 / 基因克隆 / 表达分析 {{custom_keyword}} /
winter rapeseeds / FLC transcription factor / bioinformatics analysis / gene clone / expression analysis {{custom_keyword}} /
表1 材料及来源地Table 1 Experimental materials and their origins |
| 材料名称 Material | 感温性 Temperature sensitivity | 材料来源 Origin |
|---|---|---|
| 陇油6号 Longyou 6 | 强冬性 Strong winterness | 甘肃农业大学农学院 College of Agriculture, Gansu Agricultural University |
| 陇油7号 Longyou 7 | 强冬性 Strong winterness | 甘肃农业大学农学院 College of Agriculture, Gansu Agricultural University |
| 天祝小油菜 Tianzhu small rape | 春性 Springness | 甘肃农业大学农学院 College of Agriculture, Gansu Agricultural University |
| 陇油17号 Longyou 17 | 冬性 Winterness | 甘肃农业大学农学院 College of Agriculture, Gansu Agricultural University |
| 天油2号 Tianyou 2 | 冬性 Winterness | 天水农科所 Tianshui Agricultural Science Institute |
表2 引物序列Table 2 Primer sequence |
| 引物名称 Primer | 引物序列(5'→3') Primer sequence (5'→3') | 引物用途 Usage |
|---|---|---|
| BrFLC6-F | ATGGGAAGAAGAAAAGTGGAGATCAAACG | 基因克隆 Gene clone |
| BrFLC6-R | TTACTTTAGGAGCGGGAGGGTC | |
| L6-BrFLC6-F | AGTCCAGTGCAAGTGTGAAGAAGC | 实时荧光定量PCR Real-time fluorescence quantitative PCR |
| L6-BrFLC6-R | AGTTACAGACAGAGCAGCCTTGAAC | |
| L7-BrFLC6-F | AGTCCAGTGCAAGTGTGAAGAAGC | 实时荧光定量PCR Real-time fluorescence quantitative PCR |
| L7-BrFLC6-R | AGTTACAGACAGAGCAGCCTTGAAC | |
| TZ-BrFLC6-F | AGTCCAGTGCAAGTGTGAAGAAGC | 实时荧光定量PCR Real-time fluorescence quantitative PCR |
| TZ-BrFLC6-R | AGTTACAGACAGAGCAGCCTTGAAC | |
| Actin-F | TGTGCCAATCTACGAGGGTTT | 实时荧光定量PCR Real-time fluorescence quantitative PCR |
| Actin-R | TTTCCCGCTCGGCTGTTGT |
表3 BrFLC基因家族成员基本信息Table 3 Basic information of BrFLC gene family members |
| 基因 Gene | ID | 氨基酸残基数 Amino acid | MW /kD | pI | 不稳定系数 Unstable coefficient | 亲水性 Hydrophilicity | 外显子 Exons | 亚细胞定位 Subcellular localization | 染色体定位 Chromosome location | 大白菜同源基因 The homologous in B. rapa | α螺旋 α helix /% | β折叠 β turn /% | 延伸链 Extended strand /% | 无规则卷曲 Random coil /% |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BrFLC1 | Brapa10T000598 | 197 | 22.06 | 6.01 | 39.66 | -0.44 | 7 | chlo | 2 596 272~2 601 013 | Bra009055 | 59.90 | 5.58 | 12.69 | 21.83 |
| BrFLC2 | Brapa02T000341 | 196 | 21.93 | 8.84 | 39.48 | -0.47 | 7 | Nucl | 1 679 943-~1 683 538 | Bra028599 | 65.82 | 4.59 | 12.76 | 16.84 |
| BrFLC3 | Brapa03T000416 | 197 | 21.59 | 9.23 | 40.59 | -0.29 | 7 | Nucl | 1 810 239~1 813 678 | Bra006051 | 60.41 | 4.06 | 12.18 | 23.35 |
| BrFLC4 | Brapa02T004660 | 192 | 21.50 | 6.22 | 58.14 | -0.50 | 7 | Cyto | 41 089 227~41 093 400 | Bra031888 | 59.90 | 5.21 | 11.98 | 22.92 |
| BrFLC5 | Brapa03T001557 | 196 | 22.00 | 5.53 | 42.09 | -0.39 | 7 | Nucl | 7 593 279-~7 598 575 | Bra022771 | 64.80 | 4.08 | 12.76 | 18.37 |
| BrFLC6 | Brapa02T004661 | 198 | 22.92 | 8.77 | 55.39 | -0.68 | 7 | Nucl | 41 100 098~41 104 465 | Bra031886 | 54.80 | 4.52 | 11.06 | 24.62 |
| BrFLC7 | Brapa06T002482 | 199 | 22.80 | 8.77 | 48.58 | -0.46 | 7 | Nucl | 16 449 350~16 454 026 | Bra024350 | 59.80 | 2.01 | 14.07 | 24.12 |
| BrFLC8 | Brapa06T002484 | 231 | 26.19 | 9.26 | 48.83 | -0.38 | 7 | Nucl | 16 457 667~16 461 660 | Bra024351 | 63.20 | 4.76 | 9.96 | 22.08 |
| BrFLC9 | Brapa02T004663 | 199 | 22.80 | 9.52 | 52.69 | -0.56 | 8 | Nucl | 41 109 117~41 114 125 | Bra031884 | 66.83 | 5.03 | 12.06 | 16.08 |
| BrFLC10 | Brapa10T000601 | 358 | 39.68 | 8.44 | 35.39 | -0.26 | 9 | Nucl | 2 616 921~2 624 799 | / | 49.16 | 8.38 | 18.44 | 24.02 |
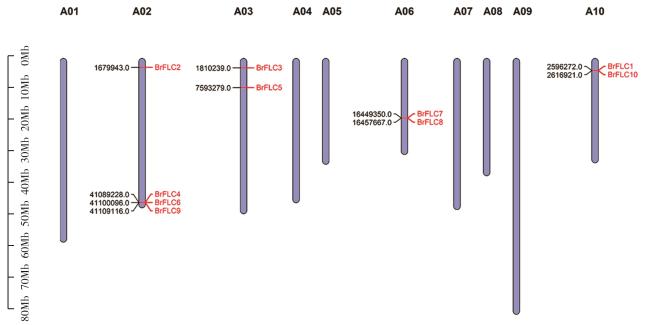
图3 白菜型冬油菜染色体上BrFLC基因的分布Fig. 3 Distribution of BrFLC genes on chromosomes of winter rapeseeds (B. rapa) |
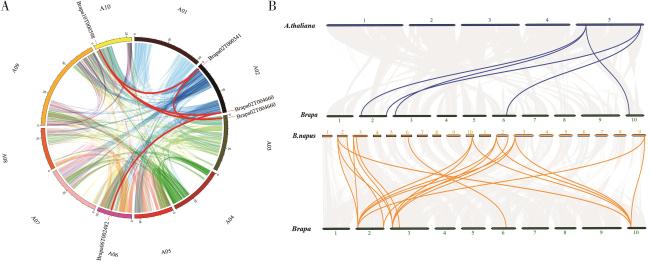
图4 白菜型冬油菜BrFLC基因家族组内共线性关系以及与拟南芥、甘蓝型油菜共线性分析Fig. 4 Analysis of collinearity in BrFLC gene family of winter rapeseeds (B. rapa) and collinearity with Arabidopsis and B. napus |

图6 BrFLC与大白菜FLC氨基酸序列比对分析Fig. 6 Amino acid sequence alignment analysis ofBrFLCs and B. rapa FLCs |
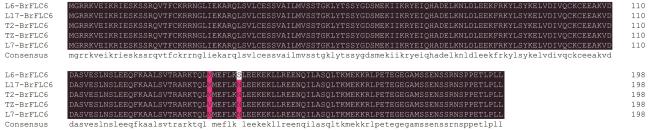
图8 不同油菜品种BrFLC6基因氨基酸序列比对分析Fig. 8 Alignment analysis of amino acid sequence of BrFLC6 gene in different rape varieties |
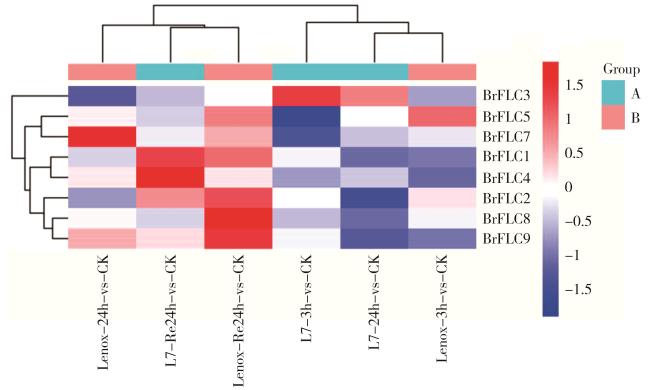
图9 4℃处理下BrFLC基因在陇油7号和Lenox生长锥中的表达模式Fig. 9 Expression pattern of BrFLC genes in Longyou 7 and Lenox growth cone at 4℃ |
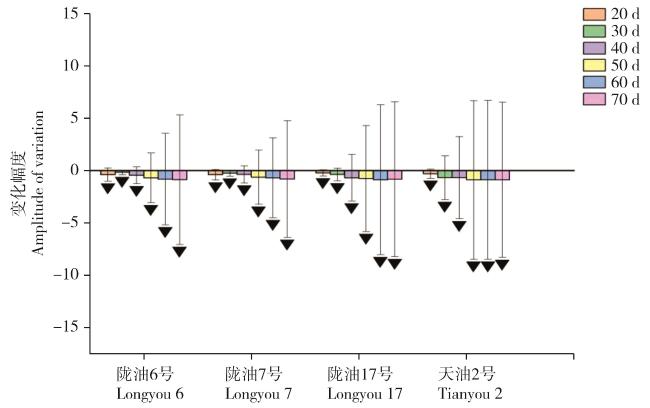
图11 春化过程中BrFLC6基因表达量幅度变化Fig. 11 Amplitude of BrFLC6 gene expression changes during vernalization |
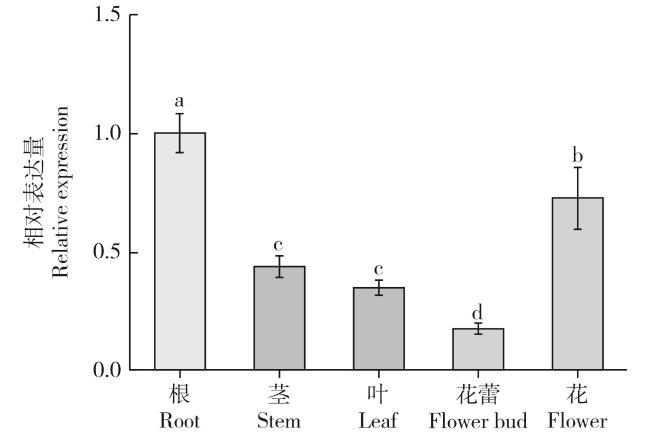
图12 BrFLC6基因在天祝小油菜不同组织中的表达Fig. 12 Expression of BrFLC6 gene in different tissues of Tianzhu small rape |
表4 不同春化时间对油菜春化率的影响Table 4 Effect of different vernalization time on rape vernalization rate |
| 时间 Time /d | 春化率 Vernalization rate | |||
|---|---|---|---|---|
| 陇油6号 Longyou 6 | 陇油7号 Longyou 7 | 陇油17号 Longyou 17 | 天油2号 Tianyou 2 | |
| CK | 0 | 0 | 0 | 0 |
| 20 | 15.6% | 13.3% | 23.2% | 46.8% |
| 30 | 26.6% | 39.6% | 46.7% | 88.9% |
| 40 | 40.0% | 46.6% | 73.3% | 98.3% |
| 50 | 73.3% | 71.6% | 93.3% | 100% |
| 60 | 86.1% | 86.6% | 96.7% | 100% |
| 70 | 95.4% | 94.3% | 97.1% | 98.3% |
| 1 |
许耀照, 孙万仓, 方彦, 等. 北方寒旱区不同抗寒性白菜型冬油菜幼苗物质转运和光合荧光动力学特征[J]. 中国油料作物学报, 2020, 42(1): 91-101. DOI:10.19802/j.issn.1007-9084.2019101 .
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 2 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 3 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 4 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 5 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 6 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 7 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 8 |
孙琦, 曲春浦, 郑美珠, 等. 小黑杨FLC基因的克隆及功能解析[J]. 植物研究, 2015, 35(3): 363-369, 377.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 9 |
王一非, 门婧婕, 张笑笑, 等. 油莎草(Cyperus esculentus) MADS-box转录因子家族鉴定及表达分析[J/OL].分子植物育种:1-15[2022-03-18].
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 10 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 11 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 12 |
蒋小刚. 甘蓝型油菜MADS-box基因家族鉴定及裂角调控网络初步分析[D]. 武汉: 华中农业大学, 2019.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 13 |
赵玉红, 马骊, 常瑜, 等. STM和CUC2基因对不同抗寒性白菜型冬油菜生长点的调控研究[J]. 中国油料作物学报, 2019, 41(2): 157-165. DOI:10.7505/j.issn.1007-9084.2019.02.001 .
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 14 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 15 |
王娟, 石大川, 陈皓宁,等. 花生高亲和硝酸盐转运蛋白基因家族生物信息学分析[J/OL]. 中国油料作物学报:1-8[2021-12-15]. DOI:10.19802/j.issn.1007-9084.2020347 .
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 16 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 17 |
马骊, 袁金海, 孙万仓, 等. 白菜型冬油菜类甜蛋白的筛选、克隆及其在低温胁迫下的表达[J]. 作物学报, 2017, 43(4): 620-628. DOI:10.3724/SP.J.1006.2017.00620 .
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 18 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 19 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 20 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 21 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 22 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 23 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 24 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 25 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 26 |
刘林波, 孙万仓, 刘自刚, 等. 白菜型冬油菜抗寒相关生理生化性状的配合力和遗传效应[J]. 中国油料作物学报, 2015, 37(6): 772-779. DOI:10.7505/j.issn.1007-9084.2015.06.006 .
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 27 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 28 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 29 |
杜春芳. 甘蓝型油菜低温诱导的转录组和蛋白组分析[D]. 武汉: 华中农业大学, 2016: 29-32.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 30 |
孙万仓, 侯献飞, 杨刚, 等. 一种利用高温诱导鉴定白菜型冬油菜抗寒性的方法, CN104982173A[P]. 2015.
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 31 |
蒲媛媛, 赵玉红, 武军艳, 等. 北方强冬性甘蓝型冬油菜品种(系)抗寒性评价[J]. 中国农业科学, 2019, 52(19): 3291-3308. DOI:10.3864/j.issn.0578-1752.2019.19.002 .
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 32 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 33 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| 34 |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
| {{custom_ref.label}} |
{{custom_citation.content}}
{{custom_citation.annotation}}
|
PDF(6450 KB)
 表1 材料及来源地表2 引物序列表3 BrFLC基因家族成员基本信息
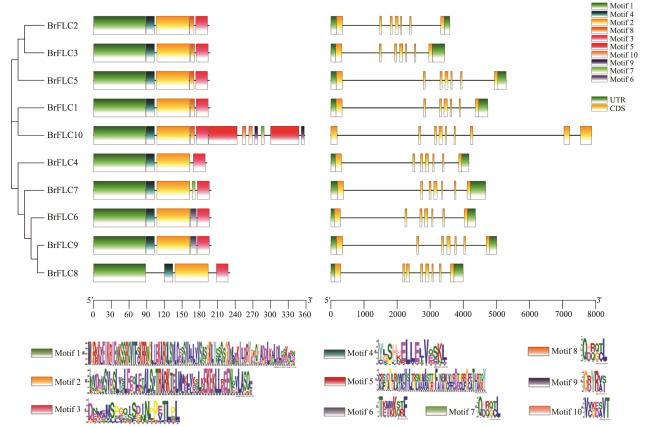
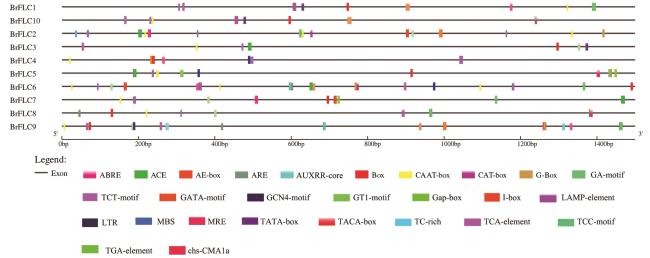
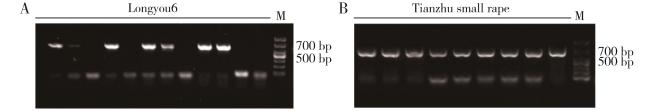
表1 材料及来源地表2 引物序列表3 BrFLC基因家族成员基本信息 图1 白菜型冬油菜、甘蓝型油菜、大白菜、拟南芥FLC基因家族进化分析图2 白菜型冬油菜BrFLC的基因结构、保守Motif分析图3 白菜型冬油菜染色体上BrFLC基因的分布图4 白菜型冬油菜BrFLC基因家族组内共线性关系以及与拟南芥、甘蓝型油菜共线性分析图5 BrFLC基因顺式作用元件分析图6 BrFLC与大白菜FLC氨基酸序列比对分析图7 BrFLC6基因菌液PCR电泳产物图8 不同油菜品种BrFLC6基因氨基酸序列比对分析图9 4℃处理下BrFLC基因在陇油7号和Lenox生长锥中的表达模式图10 春化过程中BrFLC6基因的荧光定量表达图11 春化过程中BrFLC6基因表达量幅度变化图12 BrFLC6基因在天祝小油菜不同组织中的表达表4 不同春化时间对油菜春化率的影响
图1 白菜型冬油菜、甘蓝型油菜、大白菜、拟南芥FLC基因家族进化分析图2 白菜型冬油菜BrFLC的基因结构、保守Motif分析图3 白菜型冬油菜染色体上BrFLC基因的分布图4 白菜型冬油菜BrFLC基因家族组内共线性关系以及与拟南芥、甘蓝型油菜共线性分析图5 BrFLC基因顺式作用元件分析图6 BrFLC与大白菜FLC氨基酸序列比对分析图7 BrFLC6基因菌液PCR电泳产物图8 不同油菜品种BrFLC6基因氨基酸序列比对分析图9 4℃处理下BrFLC基因在陇油7号和Lenox生长锥中的表达模式图10 春化过程中BrFLC6基因的荧光定量表达图11 春化过程中BrFLC6基因表达量幅度变化图12 BrFLC6基因在天祝小油菜不同组织中的表达表4 不同春化时间对油菜春化率的影响/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}